Differential Media:
|
|
|
|
||
Differential Media:
|
|
|
|
||
|
1. INTRODUCTION Before its recognition as a taxonomic entity, Edwardsiella tarda was shown to cause disease in humans and a variety of warm and cold-blooded vertebrates. One would think that E. tarda isolates should be relatively easy to identify in a laboratory, as they are negative for fermentation of most sugars (including lactose, sucrose, mannitol and xylose), negative for phenylalanine deamination, and positive for lysine decarboxylation and hydrogen sulfide production. As this species is a member of the Family Enterobacteriaceae and also easy to cultivate, the colonies should come up well on a selective medium such as MacConkey Agar. Furthermore, this species is known to be resistant to the antibiotic colistin, a characteristic shared by few other enteric bacteria. Keeping these characteristics in mind and remembering how the reactions from multiple substrates undergoing fermentation, deamination and decarboxylation can result in a net acidic or alkaline reaction for colonies on differential plating media (as demonstrated in the exercise here) or in such tubed media as TSI Agar and LIA, formulating a reasonably dedicated plating medium for the isolation of this organism should not prove to be difficult. If it works in theory as an appropriate selective-differential medium for E. tarda isolation, then it may very well work for culturing field samples. What follows shows how this came about. 2. DEVELOPMENT OF "ET AGAR" AND THE CHOICE OF "PRESUMPTIVE IDENTIFICATION" MEDIA The following formula was developed and given the name "ET Agar." The H2S-indicator system of XLD Agar (Difco Laboratories, Detroit, MI) was incorporated. "Fine-tuning" was accomplished with some stock cultures and field samples. Indeed, concentrations and choices of the ultimate components of a selective-differential medium are adjusted as situations arise and results dictate. A small amount of additional agar was utilized for awhile (and may show up in some references), but this further solidification of the medium was determined unnecessary. Ultimate results involving six colistin-resistant enteric genera are shown farther below.
Preparation and Use. As heating all of the ingredients together is not advised, the medium is meant to be prepared in two parts as follows for a total of one liter:
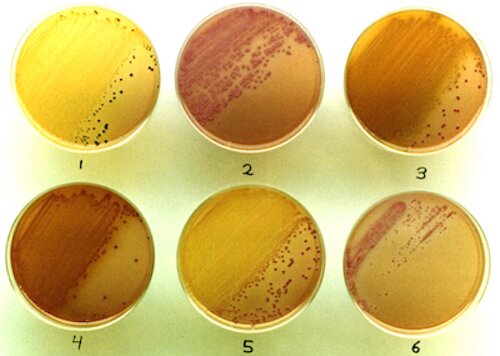
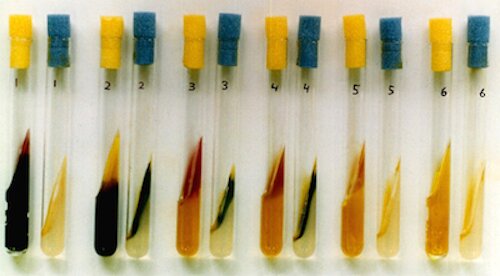
Usually, agar-containing media are initially steamed to melt the agar such that all the ingredients can be mixed thoroughly prior to autoclaving. However, I summarily decided from the outset to break tradition by not performing this pre-autoclave steaming of Mixture A, with the anticipation of doing a good mixing after the short, 10 minute autoclaving. From previous media-making experience, the mixing of the agar would be thorough enough, and overheating (however judged) should be avoided anyway. It was of great importance that Mixture A should attain plate-pouring temperature (about 50°C) before adding Mixture B, as a higher temperature might be deleterious to the sugars and other ingredients. A Required Modification of the Preparation (once the project was under way). As even some complete enteric plating media (SS Agar, for example) are not autoclaved but simply steamed to melt the agar, I was required to stop autoclaving Mixture A. This was after the project was well under way with its sampling, plating, isolating, testing, etc. Why was this so? With the advent of extreme automation in the Department's media facility, our old but reliable manual autoclave was summarily banished to the attic with other articles to eventually participate in the demolition/reconstruction project going on in that part of campus. The unexpected happy result was that the medium, now prepared with simple steaming of Mixture A (prior to the addition of Mixture B), then began to support the additional growth of a new type of colony which was similar to that of E. tarda. More about this development is discussed appropriately in the "Isolations" section below (under "Exceptions"). Reactions Expected on ET Agar and Presumptive Identification Media for Selected Colistin-Resistant Enterics. An overview of the results from the testing of representatives of six genera of colistin-resistant members of the family Enterobacteriaceae follows, but it is preceded by the following table of expected reactions on ET Agar and from substrates which could conceivably be incorporated into suitable "presumptive identification" media. Not all non-Edwardsiella species are specifically considered, and some strains of Proteus may conceivably produce black colonies resembling those of E. tarda. So, one would depend on the subsequent use of suitable screening media, and the choice was made of Phenylalanine Agar and Triple Sugar Iron Agar. The latter medium is modified with the addition of 1% mannitol which strains of E. tarda would not be expected to ferment; thus the designation "TSIM."
* = The pH is too low (especially from xylose fermentation) for the black FeS precipitate to form. Reactions Observed on ET Agar and Presumptive Identification Media for Selected Colistin-Resistant Enterics.
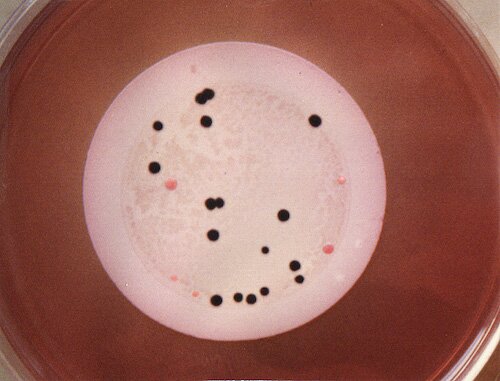
3. ISOLATIONS In 1988, I began an analysis of Midwestern stream and lake samples for Edwardsiella tarda, passing various volumes of freshly-taken and refrigerated samples (usually 1, 4, 10 and 25 ml from each sample site) through 0.45 µm membrane filters (with the use of a suction flask) which were each placed upon a plate of ET Agar. One-day incubation at 37°C was generally sufficient for good colony formation. A typical plate showing good isolation of presumptive E. tarda colonies is shown below. With exceptions noted below, (1) colonies remained black (with also a whitish, alkaline rim) after subsequent streaking on a fresh plate of ET Agar (without colistin), and (2) the presumptive identification tests for the black colonies (detailed above) turned out positive for E. tarda.
Exceptions:
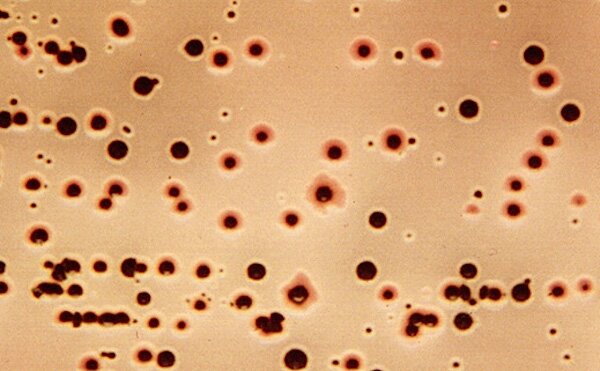
A demonstration plate on ET Agar was prepared from a mixed suspension of cells from three isolates, each representing a biogroup of E. tarda. The normal type of colony is black with a whitish rim. The CL Biogroup is seen as the tiny dark colonies, and Biogroup 1 is recognized by the red (acidic) rim due to its fermentation of sucrose and mannitol.
APPENDIX 1. SUMMARY TABLE OF THE "BIOGROUPS" AND THEIR DIFFERENTIATING REACTIONS A table showing the biochemical reactions of the several biogroups is on a convenient PDF file which can be accessed here. Included on the table is a column detailing the general reactions of the aforementioned "G30 Biogroup" which I have often characterized as "trying to pass itself off as Edwardsiella tarda" on the original filter plates. However, it is certainly different! APPENDIX 2. A NOTE FOR A BACTERIOLOGICAL JOURNAL WRITTEN IN 1991 "A Medium for the Isolation of Edwardsiella tarda" was drafted as a possible note for publication but was set aside, mainly due to the volume of work at my job as a lab instructor at U.W.-Madison. It is probably of value for its references to background material on Edwardsiella tarda. The PDF copy can be found here. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Return to the main page of |
|
Page slightly modified (for clarity) on 7/14/18 at 5:00 PM, CDT. John Lindquist: home page and complete site outline E-mail me at jlindquist 001 @ gmail.com Be sure to remove spaces (added to discourage spambots). Department of Bacteriology, U.W.-Madison |